Prof.Dr. Kaan AYDOS
DNA’nın çoğalması (replikasyonu)
Pürinler nasıl ve nereden sentezlenirler
Diyetle gelen nükleik asitlerin barsakta yıkımı
Pirimidin nukleotidlerinin yıkımı
RNA’nın sentezi (transkripsiyon)
RNA’nın transkripsiyon sonrası uğradığı değişiklikler
Translasyon işleminin özellikleri
Translasyon sonrası polipeptid zincirlerinde meydana gelen modifikasyonlar
DNA yapısında bozuklukların meydana gelmesi, bunların sonuçları ve teşhisi
Polimeraz Zincir Reaksiyonu (PCR= Polymerase chain reaction)
İnsanın varolması ve soyunu devam ettirebilmesi, hücrenin içinde, nukleusunda bulunan genetik materyalin çalışması ile gerçekleşir. Genetik bilgiler nükleik asitler içinde saklanır. İki çeşit nukleik asit vardır: DNA (deoksiribonükleik asit) ve RNA (ribonükleik asit). Organizmanın gelişmesi için gerekli bilgiler DNA içerisinde bulunurlar ve burada kodlanırlar. Burada kodlana bilgilerle hücre iki fonksiyonunu yerine getirir: çoğalma (replikasyon) ve protein sentezi. Çoğalan yani bölünen hücrenin ise özelliğine göre seçici olarak belirli bilgileri ifade edebilmesi gerekir. Hücrenin görevini yapabilmesi, sentezlediği proteinler ile diğer hücreleri veya aracı maddeleri kontrol ederek gerçekleşir.
DNA içerisinde depolanan bilgiler kodlandıktan sonra (transkripsiyon işlemi) RNA tarafından değerlendirilir. RNA bu kodlanan bilgileri sitoplazmaya taşıyarak burada protein sentezini (translasyon) yaptırır.
DNA’nın yapısı
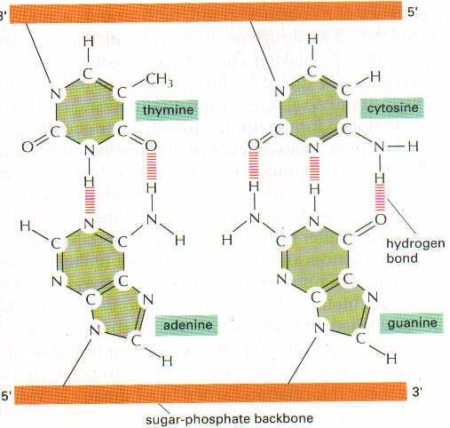
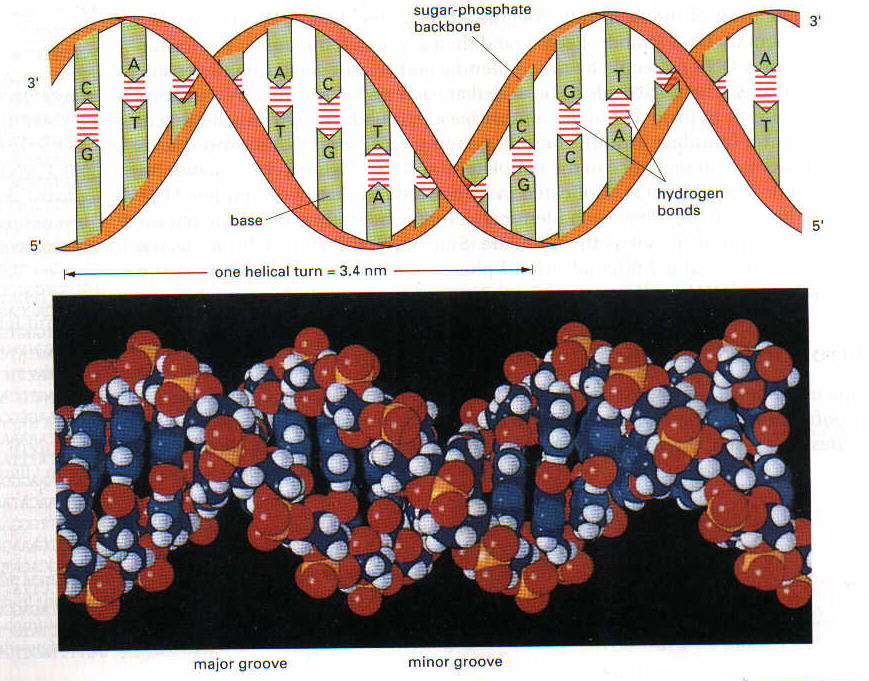 DNA bir polideoksiribonükleotid’dir, yani çok sayıda deoksiribonukleotid bir araya gelerek DNA’yı oluşturur. Nükleotidler bir pentoz şekerinin (DNA’da deoksiriboz, RNA’da bunun 2. karbonuna bağlı bir hidroksil grubu eksiktir; riboz) bir baz (pürin veya pirimidin bazlarından biri) ve bunu diğer nükleotidlere 3′-5′ fosfodiester bağı ile bağlayan bir fosfat grubundan oluşur. Fosfodiester bağı ile bir nükleotidin deoksipentozuna ait 5′-hidroksil grubu diğer nukleotidin deoksipentozuna ait 3′-hidroksil grubuna bir fosfat grubu aracılığıyla bağlanmıştır. İşte deoksiriboz-fosfat iskeleti, DNA zincirinin ana çatısını oluşturur. DNA iki zincir halinde ve iki zincir birbirine heliks yapacak tarzda sarılı bulunur. İki zincirin ortasında ise bazlar vardır. Her bir zincirdeki bazlar karşı zincirdeki sadece bir çeşit bazla birbirlerine bağlanır: Daima Adenin-Timin ile, Sitozin-Guanin ile eşleşir. Adenin ve Timin arasında 2, Guanin ve Sitozin arasında ise 3 hidrojen bağı vardır. RNA’da DNA’dan farklı olarak timin yerine urasil bulunur.
DNA bir polideoksiribonükleotid’dir, yani çok sayıda deoksiribonukleotid bir araya gelerek DNA’yı oluşturur. Nükleotidler bir pentoz şekerinin (DNA’da deoksiriboz, RNA’da bunun 2. karbonuna bağlı bir hidroksil grubu eksiktir; riboz) bir baz (pürin veya pirimidin bazlarından biri) ve bunu diğer nükleotidlere 3′-5′ fosfodiester bağı ile bağlayan bir fosfat grubundan oluşur. Fosfodiester bağı ile bir nükleotidin deoksipentozuna ait 5′-hidroksil grubu diğer nukleotidin deoksipentozuna ait 3′-hidroksil grubuna bir fosfat grubu aracılığıyla bağlanmıştır. İşte deoksiriboz-fosfat iskeleti, DNA zincirinin ana çatısını oluşturur. DNA iki zincir halinde ve iki zincir birbirine heliks yapacak tarzda sarılı bulunur. İki zincirin ortasında ise bazlar vardır. Her bir zincirdeki bazlar karşı zincirdeki sadece bir çeşit bazla birbirlerine bağlanır: Daima Adenin-Timin ile, Sitozin-Guanin ile eşleşir. Adenin ve Timin arasında 2, Guanin ve Sitozin arasında ise 3 hidrojen bağı vardır. RNA’da DNA’dan farklı olarak timin yerine urasil bulunur.
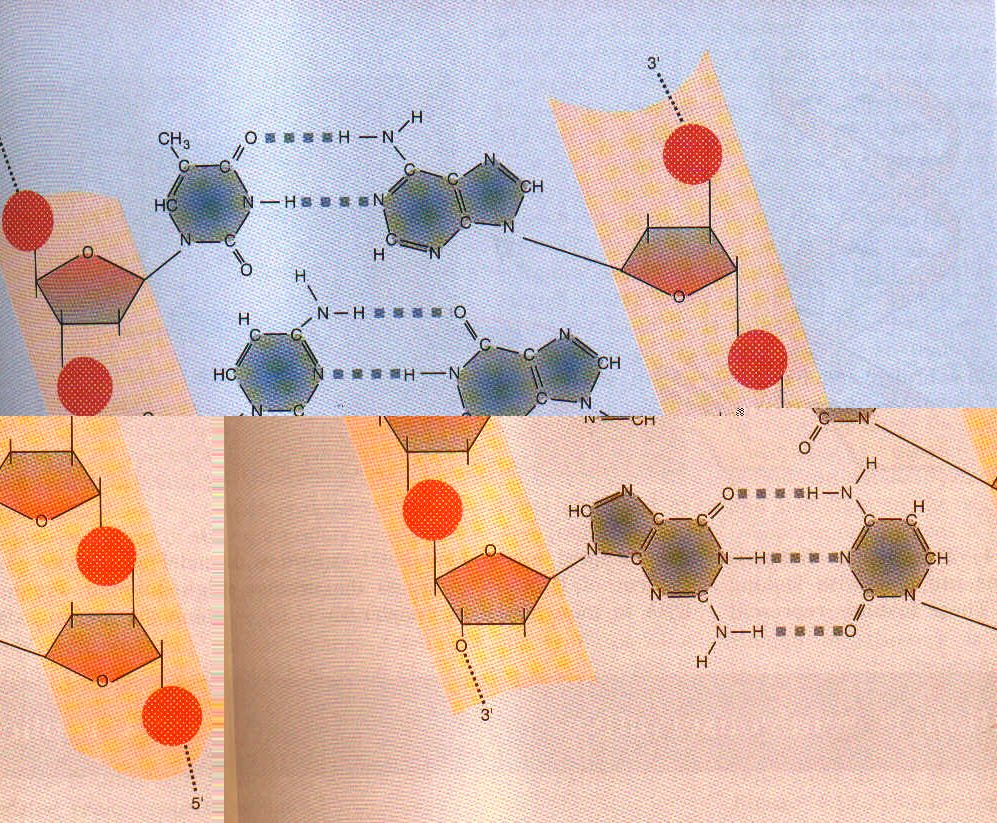
Fosfodiester bağları fosfodiesterazlar, ya da genel olarak adlandırıldığı gibi nükleazlar tarafından parçalanırlar. Bazlar arasındaki hidrojen bağları ise pH değişikliğinde veya ortamın ısısı artarsa parçalanırlar ve iki heliks denatüre olarak birbirinden ayrılır.
DNA’nın çoğalması yani replikasyonu
DNA’nın replikasyonu, iki zincirinin açılması ve her bir zincirin polimeraz enzimi vasıtasıyla kalıp olarak kullanılarak üzerine yeni bir zincirin eklenmesi şeklinde olur. Buna semikonservatif replikasyon denir. Çünkü DNA’nın iki zincirinden biri kalıp olarak kullanılır ve üzerine yeni DNA zinciri sentezlenerek yeni bir DNA oluşturulur.
DNA’nın replike olabilmesi için önce birbirine sarılmış olan DNA zincirlerinin ayrılması gerekir. Çünkü polimerazlar sadece tek DNA zincirini kalıpı olarak kabul ederler. Önce ufak bir bölgede DNA zinciri açılır. DNA replikasyonunun başladığı bu tek nukleotid dizisinin bulunduğu bölgeye “replikasyon orijini” denir. Zincir açıldığı zaman burada bir “replikasyon çatalı” oluşur. Ökaryotlarda replikasyon, DNA heliksi boyunca bir çok orijinde aynı anda başlayabilir. Replikasyon orijini daima Adenin-Timin baz çiftinin bulunduğu yerden başlar, o nedenle bunlara “konsensüs dizesi” denir.
DNA sarmalının ayrılması için önce replikasyon orijini üzerine özel proteinler bağlanır. Bu proteinlere DnaA proteinleri denir 20-50 adet DnaA proteini Adenin-Timin baz çiftinin bulunduğu replikasyon orijini üzerine bağlanarak ayrılma işlemini başlatır ve birbirine bitişik olan DNA heliksi eriyerek birbirinden ayrılmaya başlar.
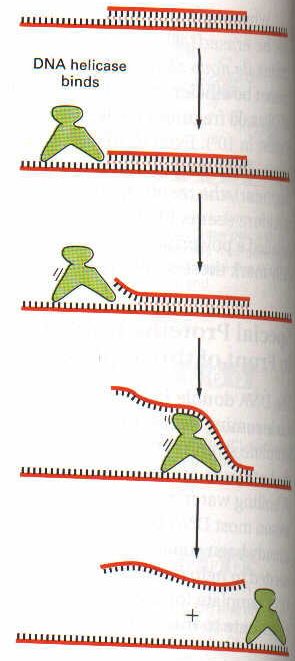
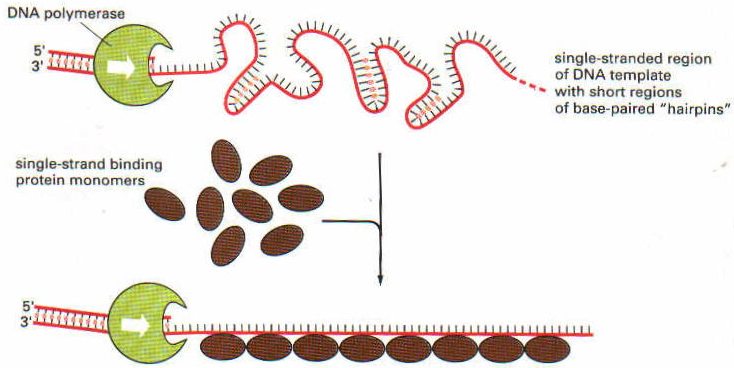
Replikasyon çatalının tam ortasına ise ayrı bir protein bağlanır: DNA helikaz. DNA helikaz, iki zincirin birbirlerinden ayrıldığı çatalın tam ortasına tutunarak zincirleri birbirinden ayrılması için zorlar. Böylece zincir ileri doğru birbirinden ayrılmış olur. Sarmallar bir kez ayrılınca, hemen TSB proteinleri gelerek ayrılmış zincirlere tutunur ve çift heliksin tekrar oluşmasına mani olurlar.
Çift heliksi oluşturan iki sarmal birbirinden ayrıldıkça, önde bulunan kromozom kısımları rotasyona uğrayarak üst üste binerler ve süperkoil’leri oluşturur. Süperkoil oluşması ise ilerideki zincirin ayrılmasını güçleştirir. DNA heliksi açılırken süperkoillerin oluşmasını bazı enzimler önler. Bunlara tip I ve tip II DNA topoizomerazları denir. Bu enzimler ayrılacak olan DNA iplikçikleri üzerinde kırılma ya da kertikler oluştururlar. Açılan bu uçlar sayesinde heliksler serbestçe dönerek, katlantılar açıldıktan sonra tekrar birbirine yapışarak zincirin bütünlüğü sağlanmış olur.
DNA zincirleri replikasyon orijininde birbirinden ayrıldıktan sonra, DNA’nın başındaki nükleotid dizisi üzerine RNA primeri gelerek yapışır. Bu primer, kalıp DNA’nın başındaki nükleotid dizisine komplementer olarak oturan, ribonükleotidlerden oluşmuş RNA parçasıdır. Primerin 3′ ucunda bulunan hidroksil grubu serbesttir. İşte DNA polimeraz bu serbest hidroksil grubuna, kalıp DNA’daki nükleotide komplementer deoksiribonükleotidi takar.
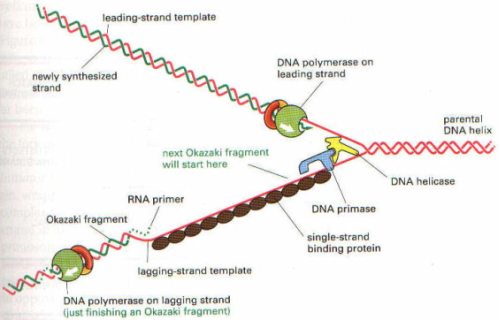
Ancak, yeni DNA zincirinin uzama yönü 5′ ucundan 3′ ucuna doğrudur. Bu durumda diğer zincirdeki ilerlemenin 3′ ucunda 5′ ucuna doğru olması gerekir. Oysa DNA polimeraz bu yönde sentez yapamaz. Bu nedenle bu zincir üzerine primaz enzimi ile birlikte bir pre-primaz protein kompleksi de bağlanır. Pre-primaz ve primaz enzimlerine birlikte primozom adı verilir. Primozomlar, açılmış DNA zinciri üzerinde bağlandıkları noktadan itibaren 5′ ucundan 3′ ucuna doğru yeni nükleotidleri sentezler, ama her nukleotid yerleştikten sonra primozom geri doğru giderek, diğer zincirle aynı yönde, yani 5′ ucundan 3′ ucuna doğru sentezi gerçekleştirmiş olur.
DNA sentezini gerçekleştiren enzim DNA polimeraz III’dür. Sentez işlemi, önünde yeni bir RNA primeri takılı bölgeye gelene kadar devam eder. Primere rastlayınca, DNA polimeraz I enzimi bu RNA’yı ayırır ve yerine yeni bir DNA nükleotidi takar. Bu nükleotidin, önünde daha önce oluşmuş nükleotide bağlanmasını DNA ligaz enzimi gerçekleştirir. Böylece sadece DNA’lardan oluşmuş zincir devamlı sentezlenmiş olur.
Bu şekilde, DNA çift heliksini oluşturan iki zincir birbirinden ayrılmış ve üzerlerine yeni bir zincir eklenerek, iki yeni DNA çift zincirli heliksi meydana gelmiş olur.
İnsan hücresinde 46 adet kromozom vardır. Kromozomların toplam uzunluğu yaklaşık 1 metredir. Hepsi nukleus içerisinde paketlenmiştir. DNA replikasyon işlemini tek başına yapamaz. Başka proteinlerle ilişki içinde bulunur. DNA ve proteinler özel bir şekilde bir araya paketlenmiştir. Bu proteinlere histon denir. DNA histonlara sıkıca tutunarak nukleozom’ları oluşturur. Nukleozomlar bir iplik üzerinde dizilmiş tespih taneleri gibi gözükür. İki nukleozom arasında 50 nükleotidlik bir bağlayıcı DNA parçası bulunur. Yani DNA histonlara (5 adet histon vardır: H1, H2A, H2B, H3 ve H4; DNA sadece H1 dışındaki histonlara sarılır, H1’in başka görevi vardır) sarılmış, kümeler halinde görülür ve bu kümeler birbirine bağlayıcı DNA dizileri ile birleşmişlerdir. DNA replikasyonu sırasında aynı anda yeni histonlar da sentezlenirler.
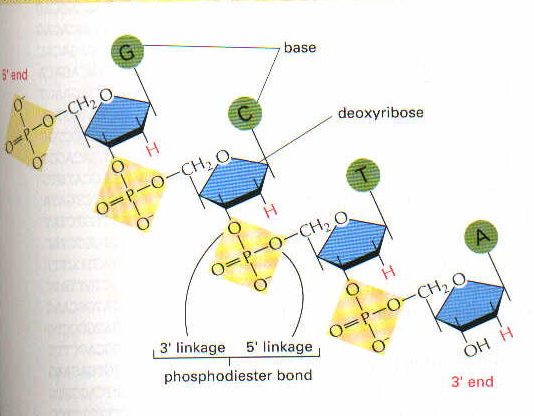
DNA’nın yapısında nukleotidler bulunur. Nukleotidler ise bir baz, şeker olarak bir pentoz monosakkaridi, ve fosfat grubundan meydana gelir. Bu şeker eğer riboz ise ribonukleozid (RNA’da) deoksiriboz ise deoksiribonukleozid (DNA’da olduğu gibi) meydana gelmiş olur (ribozun 2. karbonunda hidroksil grubu yoksa buna deoksiriboz denir).
Bazlar pürin veya pirimidin olmak üzere 2 grup halindedir. Pürin bazları hem DNA hem de RNA’nın yapısında bulunurlar: adenin ve guanin. Pirimidinlerden ise sitozin hem DNA hem de RNA’da bulunurken, urasil sadece RNA’da timin ise sadece DNA’da bulunur. Timin’de bir metil grubu varken, urasil’de yoktur.
Bir baz ile pentoz şekeri birleşince bir nukleozid denir. Buna fosfat grubu eklenince nukleotid adını alır. Fosfatlar pentozun 5′ ucundaki karbon atomuna bağlı OH grubuna bağlanırlar. Aslında fosfat bağları iki nukleotidin birbirine bağlanmasında rol alırlar ve bu nedenle bir nukleotidin 5′ ucundaki OH grubuna diğer nukleotidde ise 3′ ucundaki OH grubuna bağlanarak, iki nukleotidi birleştirmiş olurlar.
Eğer pentozun 5′ ucuna 1 adet fosfat grubu eklenmişse nukleotid monofosfat (örneğin AMP), 2 adet fosfat grubu eklenmişse nukleotid difosfat (örneğin ADP), 3 adet fosfat grubu eklenmiş ise nukleotid trifosfat (ATP) adı verilir.
Pürinler nasıl ve nereden sentezlenirler
Pürin bazını 9 adet atom oluşturur. Bu atomların her biri ise değişik amino asitlerden (aspartik asit, glisin ve glutamin) ve CO2 ile tetrahidrofolat gibi bileşiklerden gelir. Yani aslında DNA’yı oluşturan nukleotidler, ortamda hazır bulunan bir çok değişik bileşiklerden sentezlenmektedir.
Nukleotidler sentezlenirken önce riboz şekeri kullanılarak ribonukleotidler meydana getirilir daha sonra bundan deoksiribonukleotidler yapılır.
Sentezleme işlemi daha önce değişik reaksiyonlarla oluşmuş riboz-5-fosfat’dan PRPP (fosforibozil pirofosfat)’ın meydana gelmesiyle başlar.
PRPP’ye ise glutamin eklenerek fosforibozilamin sentezlenir.
Buna da yukarıda sayılan glisin eklenerek ve folat ile CO2’in katalizörlüğü etkisiyle İMP (inozin monofosfat) meydana gelir.
İMP’dan ise sonuçta AMP ve GMP nukleotidleri oluşur.
AMP ve GMP nukleozid monofosfatlardır (nukleozid monofosfat demek, yapısında bir fosfat grubu taşıyan nukleotid ile aynı anlama gelir). Bunlara ATP’den fosfatların eklenmesi sonucu ADP ve GDP ile ATP ve GTP’lar meydana gelir. Enzim adenilat (guenilat) kinazlardır.
Diğer yandan, hücre içinde bulunan nükleik asitlerin yıkımından ya da diyet ile gelen pürinlerden de tekrar pürinler sentezlenebilirler (yan yol).
Pürinler yıkılınca sonuçta ürik asit oluşur. Ürik asitte idrar ile atılır.
Yıkım sırasında sırasıyla AMP, Adenozin, İnozin, Hipoksantin, Ksantin ve Ürik asit meydana gelir. AMP aynı zamanda İMP’ye dönüşerek de yıkılabilir.
GMP’dan ise sırasıyla Guanozin, Guanin, Ksantin ve Ürik asit meydana gelir.
İMP ise sırasıyla İnozin, Hipoksantin, Ksantin ve Ürik asit’e yıkılır.
Diyetle gelen nükleik asitlerin barsakta yıkılımı
Besinlerin içerisinde bulunan RNA ve DNA, midede düşük pH’da denatüre olur ve önce pankreastan salgılanan nukleaz enzimleri ile oligonukleotidlere ayrışırlar. Bunlarda yine pankreastan gelen fosfodiesterazlarla daha ileri hidrolize uğrayarak mononukleotidleri meydana getirir. Mononukleotidlerde, nukleotidazlarla nukleozidlere, bunlarda nukleozidazlarla nukleozidlere ayrıştırılır. Nukleozidler barsak epiteli mukoza hücreleri tarafından absorbe edilirler. Ya da nukleozidler önce yıkılarak serbest bazlar açığa çıkar, sonra bu bazlar barsaktan emilirler.
Emilen bazlar ya ince barsak mukoza hücreleri içerisinde ürik asite dönüştürülerek idrarla atılır, ya da kan dolaşımına geçerler. Bunlar çok az olarak doku nukleik asitlerinin sentezinde kullanılırlar, genelde barsak florası tarafından metabolize edilirler.
Pirimidin bazlarını oluşturan karbon ve azot atomları glutamin, CO2 ve aspartik asitten gelir.
Önce glutamin ve CO2 birleşerek karbamoil fosfatı yapar.
Bundanda bir dizi reaksiyon neticesi orotik asit meydana gelir.
Orotik asite, PRPP’dan gelen riboz-5-fosfat eklenerek orotidin monofosfat (OMP) sentezlenir. Bundan da bir karboksil grubu uzaklaşarak, üridin monofosfat (UMP) meydana gelir.
UMP, ATP’den aldığı fosfat gruplarıyla UDP ve UTP’a dönüşür. UTP’nin glutamin ile birleşerek aminasyonu neticesi ise stidin mono-di-trifosfat (CMP, CDP, CTP) meydana gelir.
Pirimidin nukleotidlerinin yıkımı
Pirimidin halkası önce açılır ve beta-alanin ile beta-aminoizobütirat meydana gelir. Beta-alaninden asetil-CoA, beta-aminoizobütirattan ise süksinil-CoA oluşur.
Ya da, pirimidinler yan yollara girerek nukleotidlere dönüşebilirler.
Nukleotid sentezinde önce ribonukleotidler (nukleotid difosfatlar yani ADP, GDP, CDP, UDP) sentezlenir ve bunlardan ribonukleotid redüktaz enzimi vasıtasıyla deoksiribonukleotidler (dADP, dGDP, dCDP, dUDP) yapılır.
Timidin DNA’da vardır ama RNA’da bulunmaz. Timidin monofosfat (TMP) dUMP’dan sentezlenir. Urasilden farklı olarak timidinde metil grubu bulunur. Zaten dUMP’da tetrahidrofolat’tan bir metil grubu alarak TMP’ı yapar.
RNA (RiboNukleik Asit) sentez ve fonksiyonları
Bir organizmanın genetik yapısını DNA üzerinde bulunan deoksiribonukleotid dizeleri belirler. Ama bu genetik yapının fonksiyon görebilmesi için, yani ifade edilebilmesi için, ribonukleik asitler (RNA) gereklidir. Önce DNA üzerindeki genetik bilgiler, RNA polimeraz enzimi aracılığıyla RNA molekülleri üzerine kopyalanır (transkripsiyon işlemi). Bu RNA’lara haberci RNA (mRNA) denir. mRNA’lar genetik bilgileri sitoplazmaya taşırlar ve burada ribozomların üzerinde yerleşirler. Ribozomlar ise ribozomal RNA (rRNA)’lardan meydana gelmişlerdir. Ribozomlar üzerine yerleşen mRNA’lardaki genetik bilgilere uygun olarak, sitoplazmada bulunan aminoasitler, taşıyıcı RNA (tRNA) tarafından alınarak mRNA üzerindeki uygun yerlere yerleştirilirler. Daha sonra bu aminoasitler birbirleri ile birleşerek proteinleri yapar (translasyon işlemi).
DNA üzerinde RNA sentezinin nerede ve ne sıklıkla olacağı, nereye kadar süreceği DNA dizelerinde bulunan bazı sinyaller tarafından kararlaştırılır. Bir hücrede bulunan DNA üzerindeki bazı genler transkripsiyona uğrayarak ifade edilebilirlerken, aynı genler başka bir hücrede ifade edilmeyebilirler. İşte bu nedenle farklı cins hücrelerin fonksiyonları da farklıdır, yani hücrelerin fonksiyonel olarak birbirinden farklı olması, DNA’daki transkripsiyon olayının farklılığına bağlıdır.
RNA’lar da DNA’lar gibi birbirlerine fosfodiester bağları ile bağlanmış mononukleotidlerden meydana gelmişlerdir. Ama şeker olarak deoksiriboz yerine riboz, baz olarak ise timin yerine urasil içerirler.
Bunlar sitoplazmada ribozomlarda bulunurlar. Ribozomlar protein sentezinin olduğu yerdir. Ökaryotik hücrelerin içerisinde 4 farklı büyüklükte rRNA bulunur: 28S, 18S, 5.8S ve 5S. Hücredeki RNA’ların %80’i rRNA’lardır.
Not: S birimi (Svedberg), bileşiğin molekül ağırlığı ile ilgili bir birimdir.
4S büyüklüğündedir. 74-95 nukleotidden meydana gelmiştir. Sitoplazmada bulunan aminoasitlerin mRNA üzerine taşınmasında görev alırlar. Vücutta bulunan proteinlerin yapısında 20 adet aminoasit bulunur. Her bir aminoasit için ayrı, özgün bir tRNA vardır. Her tRNA kendine ait aminoasiti taşır. tRNA’nın 3′ ucunda bir CCA dizesi bulunur. Aminoasit buraya bağlanır. Sonra bu tRNA, kendine bağlanmış olan aminoasiti ribozoma götürür ve burada mRNA üzerindeki genetik koda uygun yere yerleşir. tRNA’nın mRNA üzerine yerleşen kısmına antikodon denir. tRNA mRNA üzerine bağlandıktan sonra, taşıdığı aminoasit yanındaki aminoasit ile birleşerek, polipeptid zinciri uzar.
Tüm RNA’ların %15’ini oluşturur.
Molekül ağırlıkları çok farklıdır. DNA’dan aldığı genetik bilgiyi, sitoplazmadaki ribozomlara taşır. Burada protein sentezi için bir kalıp görevi yapar. Tüm RNA’ların %5’ini oluşturur. mRNA DNA’dan ilk sentezlendikten sonra bazı modifikasyonlara uğrayarak, 3′ ucuna çok sayıda adenin nukleotidinden oluşmuş poli-A kuyruğu bağlanır, 5′ ucuna ise trifosfat bağı ile bir 7-metilguanozin bağlanır.
RNA’nın sentezi, transkripsiyon
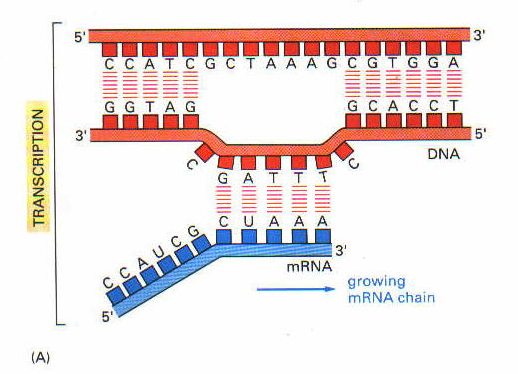
RNA’nın sentezlenmesi için önce DNA çift ipliği gevşeyerek birbirinden ayrılır. Bundan sonra RNA polimeraz ve bazı transkripsiyon faktörleri (bu faktörler promoter bölgesinde veya yakınındaki, RNA polimerazın bağlandıklarından farklı nukleotid dizelerine bağlanırlar) DNA’daki özgün dizeleri tanıyarak buralara bağlanırlar. Bağlanma işlemini takiben transkripsiyon da başlar.
DNA’da transkripsiyona uğrayacak gen kısmının başında bulunan nukleotid dizesine promoter bölgesi adı verilir. RNA polimeraz işte buraya bağlanır. Sonra DNA’nın bu kısmını kalıp olarak kullanarak buna komplementer bir RNA oluşturur (mRNA). DNA üzerindeki bazı belirli sonlanma bölgelerine gelince, RNA polimeraz transkripsiyonu durdurur.
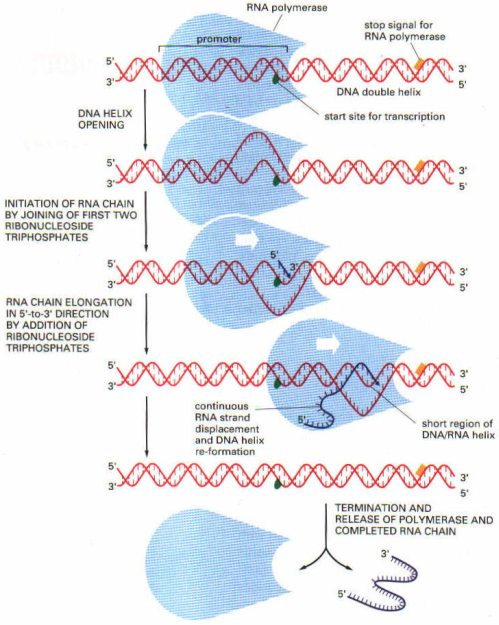
RNA’nın sentezi, RNA’nın 5′ ucundan 3′ ucuna doğru olur. DNA kalıbının kopyalanarak yeni bir RNA oluşması işlemi aynı DNA’daki gibidir. Tek farkı adenin bazının timin yerine urasil ile eşleşmesidir.
DNA üzerine RNA polimeraz enzimi bağlanınca, DNA heliksinin o kısmı lokal olarak ters yönde dönerek açılır. Ancak, enzimin önünde ve arkasında, sarmalların üst üste birikmeleri netcesi bazı süperkoiller de oluşmaktadır. Bu süperkoiller topoizomeraz I (negatif süperkoillerin açılması için) ve giraz (pozitif süperkoillerin gevşemesi için) enzimlerinin buraya bağlanarak koparma-yapıştırma işlemleri yapmasıyla gevşer ve açılırlar.
RNA sentezinin sonlanması iki şekilde olabilir: 1) DNA üzerinde bazı sonlanma bölgeleri vardır ve RNA polimeraz buraya gelince bu nukleotidleri tanıyarak işlemi sonlandırır. 2) P-faktörü denilen (ATPaz aktivitesi vardır) protein yapısında bir sonlanma faktörü vardır ve sentezlenen RNA’nın DNA kalıbından ve RNA polimeraz zincirinden ayrılmasını ve salınmasını sağlar. P’den bağımsız sonlanmada, sentezlenen RNA ipliği sona yaklaştıkça kendi üzerine kıvrılır ve daha fazla ilerlemesine mani olur. Bu kıvrımın yapısında bulunan guanin ve sitozin arasındaki 3’lü hidrojen bağları, güçlü bir yapı oluşturarak, kıvrımın dayanıklılığı sağlanır. Kıvrımın hemen yakınında ise bol miktarda urasil bazı bulunur. Urasil bazları ise adenin ile zayıf 2’li hidrojen bağları yaptıkları için, bu RNA dizesinin kalıptan kolayca ayrılmasını sağlar.
Promoter bölgesinden sonlama bölgesine kadar uzanan DNA kısmına “transkripsiyon birimi” denir. RNA polimeraz tarafından sentezlenen yeni ürüne ise “primer transkript” adı verilir. Primer transkriptler daha sonra post-transkripsiyonel değişikliklere uğrarlar ve fonksiyonel özgünlükte RNA haline gelirler.
RNA polimerazın ilk bağlandığı DNA dizeleridir. Transkripsiyonu yapılacak genin genellikle başında bulunur. mRNA’nın ilk bazının kodlandığı nukleotid dizelerinin bulunduğu yer transkripsiyonun başladığı yerdir. Promoter bölgesi ise bu noktadan itibaren geriye doğru uzanan çok sayıda nukleotidden meydana gelmiştir. Transkripsiyona başlanan yerden 25 baz geride özel bir nukleotid dizesi vardır: Hogness box denilir ve TATA (timin-adenin-timin-adenin) şeklinde 4 nukleotidden oluşmuştur. Bunun da gerisinde, yani başlangıç nukleotidinden yaklaşık 70-80 baz geride bir diğer özel bölge bulunur ki bunada CAAT box adı verilir.
İşte, promoter bölgesinde bulunan bu özelleşmiş nukleotid dizeleri, RNA polimeraz ve transkripsiyon faktörleri için tanınma bölgeleri olarak görev yaparlar. Yani RNA polimeraz ve transkripsiyon faktörleri önce gelerek buraya bağlanırlar. Ancak, transkripsiyon faktörleri özgün DNA dizelerine bağlandıkları gibi, RNA polimeraza veya kendilerine de bağlanabilirler. Bir de DNA üzerinde “hızlandırıcılar” denilen ve RNA polimerazın transkripsiyona başlama hızını arttıran özel nukleotid dizeleri vardır. Bunlar transkripsiyon başlangıç noktasından önce veya sonra yerleşmiş olabilirler. Lokalizasyonları çok uzaklarda da bulunabilir.
Ökaryotlarda 3 tip RNA polimeraz vardır. RNA polimeraz I rRNA prekürsörlerini, polimeraz II mRNA ve küçük nükleer RNA (snRNA)’ları, polimeraz III ise tRNA’ları ve bazı küçük rRNA’lar ile snRNA’ları sentezlerler.
RNA’nın transkripsiyon sonrası uğradığı değişiklikler
DNA’dan sentezlenen ilk RNA dizesi, yani primer transkript, DNA’nın özgün bir kopyasıdır. Bu RNA’ların daha sonra bazı işlemler geçirmesi gerekmektedir.
rRNA’lar sentezlendikten sonra RNAaz denilen enzimler ile daha küçük parçalara ayrılırlar ve neticede 28S, 18S, ve 5.8S’lik rRNA’lar meydana gelir. Bunlar da ayrıca bazı proteinlerle etkileşime uğrarlar.
tRNA’lar ise yine aynı enzimle belirli yerlerinden kırılarak daha küçük molekülleri meydana gelir. 3′ ucuna da nukleotidiltransferaz enzimi vasıtasıyla bir CCA dizesi eklenir. Bu uca aminoasitler bağlanacaktır. tRNA’nın antikodon bölümü ise mRNA üzerinde kendine uygun nukleuotid dizesine bağlanmaktadır.
mRNA ise polimeraz II enzimi tarafından nukleusta sentezlendikten sonra (heterojen nukleer RNA, hnRNA) 3 önemli değişime uğrar. 1) 5′ ucuna 7-metil guanozintrifosfat eklenir. Bunun guanozintrifosfatını guaniltransferaz enzimi takar. Metil grubu ise S-adenozil metioninden gelir ve metiltransferaz enzimi vasıtasıyla buraya eklenir. 2) 3′ ucuna 40-200 adet adenin nukleotidi eklenir (poli-A kuyruğu). Bu poli-A kuyruğu DNA’dan gelmez. Transkripsiyon işlemi tamamlandıktan sonra poli-A polimeraz enzimi tarafından yapılır. mRNA’ya bu kuyruğun takılması için sinyal, DNA üzerindeki poliadenilasyon sinyal dizesi tarafından gönderilir. 3) Son olarak, mRNA’nın yapısında bulunan ve protein kodlamayan intron kısımları çıkarılarak, protein kodlama özelliği taşıyan eksonlar tekrar uç uca birleştirilirler. Bu işlemi küçük nukleer ribonukleoproteinler (snRNA) gerçekleştirir.
Ancak yukarıda belirtilen modifikasyonlar yapıldıktan sonra RNA’lar sağlıklı fonksiyon yapabilir hale gelirler. RNA’ların fonksiyon yapabilmeleri için sadece DNA’dan sentezlenmeleri yetmemekte, çevrede bulunan diğer proteinler ya da enzimlerle de etkileşime uğramaları gerekmektedir.
Kromozomlarda DNA içinde depolanan genetik bilgi, DNA’nın replikasyonu ile yavru hücrelere iletilir. Bu hücrenin bu genetik bilgiyi ifade edebilmesi için ise bir peptid, ya da protein, sentezlemesi gerekir. İşte bu polipeptid zincirinin, DNA’dan oluşabilmesi için de önce transkripsiyon ile mRNA sentezlenmesi gerekir. mRNA, DNA’dan aldığı genetik kopyayı sitoplazmadaki ribozomlara götürür, ribozomlardada tRNA’ların getirdiği aminoasitler yan yana dizilerek, polipeptid zinciri meydana gelmiş olur.
Hangi aminoasitlerin seçileceği, mRNA üzerinde bulunan genetik kod ile belirlenmiştir. Genetik kod ise nukleik asit dizelerinden, nukleotidlerden oluşmuştur. Bu nukleotidlerin dizilişine göre de proteinleri oluşturan aminoasitlerin seçimi yapılır. Böylece bir protein sentezlenmiş olur.
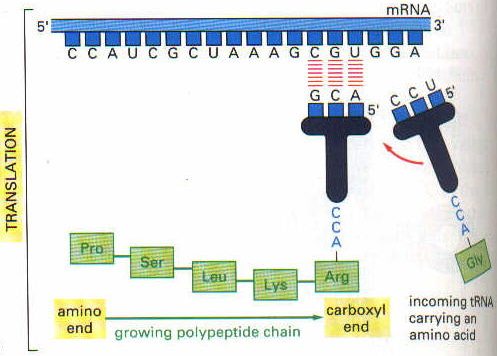
Nukleotidlerin dizilmesinde bir bozukluk olursa, neticede polipeptid zincirine yanlış bir aminoasitin bağlanmasına yol açacak, bu da hastalığa neden olabilecektir.
Genetik kod, bir nukleotid baz dizesinin karşılık geldiği aminoasit dizesini belirtir. Üç tane nukleotid bazı bir kodon oluşturur. Her bir kodon da bir aminoasiti gösterir. Ne kadar kodon varsa, o kadar uzunlukta protein sentezlenir. Yani, kodonlar hep birlikte genetik kodu tayin ederler.
Kodonları mRNA üzerinde bulunan adenin, guanin, sitozin ve urasil bazları oluşturur. Her kodonda bu bazlardan 3 tanesi bulunur. Kodonları yazarken, mRNA”ın 5′ ucundan 3′ ucuna doğru yazılır. Toplam 64 değişik kodon vardır. Vücutta 20 aminoasit bulunur. Demekki, aynı aminoasit birden fazla kodon tarafından da kodlanabilmektedir.
Genetik kodun tercüme edilerek protein sentezlenmesi işlemine translasyon denir. Örneğin CAU kodonu histidini kodlar. AUG ise metionin aminoasitini kodlar. Bu aminoasitlerde yan yana gelerek polipeptid zincirini, yani proteini oluşturur.
mRNA’lar ribozomlara gelip, üzerindeki kodonlara uygun aminoasit seçimi yapılırken, bazı kodonlara gelince bu sentez işlemi sona erer. Yani protein sentezinin bittiğini gösteren bazı kodonlar vardır. Bunlara sonlanma kodonları adı verilir. UAG, UGA ve UAA böyle sonlanma kodonlarıdır ve herhangi bir aminoasit kodlamazlar. Sadece sentezin bittiğine işaret ederler.
Translasyon işleminin özellikleri
Bir proteinin sentezlenebilmesi için öncelikle bunu oluşturacak aminoasitlerin, aminoasitleri kodlayan ve taşıyan mRNA ve tRNA’nın, üzerinde polipeptid zincirinin oluşturulduğu ribozomların, bütün bu işlemler sürecinde kullanılan protein faktörlerin, enzimlerin ve enerji kaynaklarının sitoplazma içinde bulunması gerekmektedir.
Amino asitler: Sentezlenecek olan proteinin yapısında bulunan amino asitlerin hepsinin bu sırada ortamda hazır bulunmaları gerekmektedir. Eğer diyette bu amino asit az bulunuyor veya yoksa, o zaman eksik olan amino asiti kodlayan yerde tRNA bunu bulamıyacağı için, protein zinciri sentezi o noktada aksar. Bu nedenle diyette esansiyel amino asitlerin mutlaka bulunması zorunluluğu vardır.
tRNA: mRNA’daki şifreye uygun olarak sitoplazmadan amino asitleri alarak ribozomlara taşır. İnsanda en az 50 çeşit tRNA bulunmaktadır. Ama bunların taşıyacağı sadece 20 amino asit mevcuttur. Çünkü bir amino asite ait birden fazla tRNA bulunur. 3′ ucunda amino asitin tutunduğu CCA kodonu bulunur. Antikodon denen özel bir üçlü nukleotid dizesi içerirler. Bu anti kodon, mRNA üzerinde bulunan kendine ait kodonu tanır ve buraya tutunur. İşte bu tutunma noktasında, tRNA üzerindeki amino asit peptid zincirine katılır.
mRNA: Protein sentezinde mRNA bir kalıp olarak kullanılır. Üzerinde DNA’dan aldığı şifreler bulunur. Bu şifrelere, yani kod’lara, uygun amino asitleri ise tRNA sitoplazmadan toplayarak getirir, ribozomlar içinde bu amino asitler yan yana eklenerek polipeptid zinciri, protein, oluşmuş olur.
Amino asit-tRNA sentetazlar: Amino asitlerin kendi tRNA’larına 3′ uçlarından bağlanmalarını sağlar.
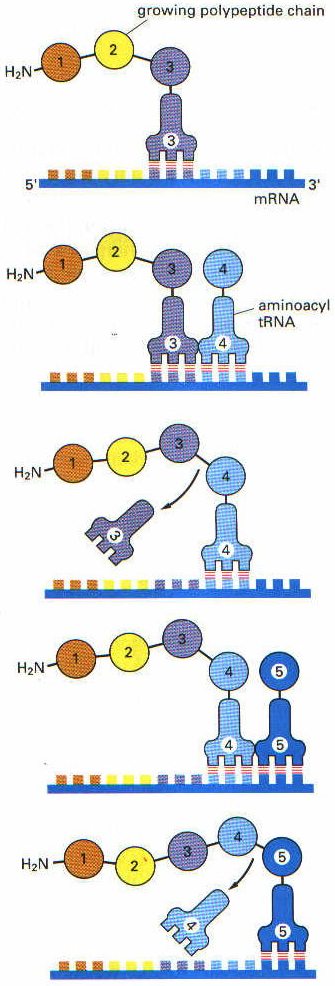
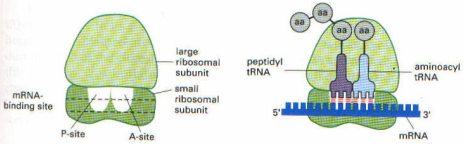
Ribozomlar: Protein ve rRNA’lardan oluşmuşlardır. Protein sentezi burada gerçekleşir. Biri büyük diğeri ise küçük iki alt birimden oluşmuşlardır. Alt birimde ise A ve P olmak üzere iki bölge bulunur. Bu bölgelere tRNA bağlanır. A ve P bölgeleri aynı zamanda mRNA’nın da peş peşe iki kodonunu kaplar. mRNA, A bölgesi içinden geçerken, mRNA’nın burada bulunan kodonuna uygun tRNA sitoplazmadan ilgili amino asiti alarak buraya gelir ve kodonun üzerine oturur. Amino asitler birbirleri ile birleşerek peptid zincirini oluşturur. Oluşan peptid zincirini taşıyan tRNA ise artık ribozomun P bölgesine geçer.
Protein faktörler: Protein sentezinde rol alan başlangıç, uzama ve sonlanma (salınım) faktörleridir.
ATP ve GTP enerji kaynakları: Polipeptid zinciri sentezlenirken enerji ATP ve GTP’lerin hidrolizlerinden sağlanılır.
tRNA amino asiti ile birlikte, kendi anti kodonuna uyan nukleotid dizesi içeren mRNA kodonu üzerine bağlanır. Bazı amino asitler birden fazla kodon ile kodlanmıştır. Bu durumda bu amino asite özgün tRNA’nın birden fazla kodonu tanıması gerekir. Bu tanıma işlemi “wobble hipotezi” ile açıklanmaktadır.
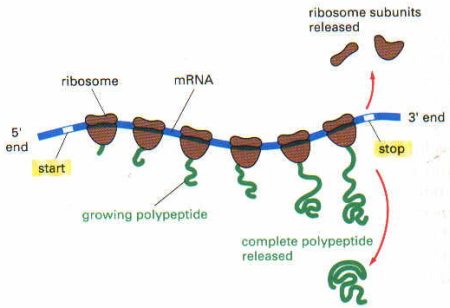
Protein sentezi yani translasyon aynı zamanda tercüme anlamına da gelir, çünkü protein sentezi esnasında nukleotid alfabesi amino asit alfabesine çevrilmektedir. mRNA’da 5′ ucundan başlayarak 3′ ucuna doğru nukleotid dizeleri okunurken, buna paralel olarakda protein sentezi amino ucundan karboksil ucuna doğru ilerler. İnsanda bir mRNA zinciri sadece bir tane peptid zincirini yapabilir.
Protein sentezinin 3 aşaması vardır:
Önce mRNA gelerek ribozomun bir alt birimindeki, kendine uyan rRNA’sına bağlanır. mRNA üzerinde bu bağlanmanın gerçekleştiği özel bir bölge bulunur. mRNA üzerinde ilk amino asiti kodlayan nukleotid dizesi AUG yapısındadır. İşte bu kodondan biraz daha 5′ ucuna yakın bir lokalizasyonda bu bağlanmayı gerçekleştirecek nukleotid dizesi vardır. Bu bağlanma bölgesi nukleotidleri, ribozomun alt biriminde rRNA’nın 3′ ucuna yakın bir loklaizasyondaki, kendine komplementer bir nukleotid dizesine bağlanır. Yani, mRNA’nın 5′ ucu ile, rRNA’nın 3′ ucu arasında komplementer bir baz eşleşmesi gerçekleşir.
Bütün bu bağlanma ve başlangıç işlemleri protein yapısındaki özel faktörler tarafından hızlandırılır.
Translasyon mRNA’nın 5′ ucundan başlar ve ribozom mRNA molekülü üzerinde kayarak ilerler. Bu sırada mRNA üzerinde ribozom 5′ ucundan 3′ ucuna doğru, her seferinde 3 nukleotid yani bir kodon atlayarak ilerler (translokasyon). İlerleme sırasında mRNA’nın hangi kodonu üzerine geldiyse, buraya uyan amino asiti taşıyan tRNA gelir ve bağlanır. tRNA’nın üzerindeki amino asit ise kendinden önce oluşmuş polipeptid zincirinin karboksil ucuna eklenir. Ribozom mRNA üzerinde hareket ettikçe, amino asitlerin birbirine eklenmesiyle polipeptid zinciri de uzamaya devam eder. Uzama faktörleri bu işlemi stimüle ederek hızlandırır. Peptid bağlarının oluşmasını peptidil transferaz enzimi sağlar. Bu enzim rRNA’nın yapısında bulunur. Üzerindeki amino asit peptid zincirine eklenmesiyle yüksüz hale gelen tRNA ise tekrar sitoplazmaya salınır.
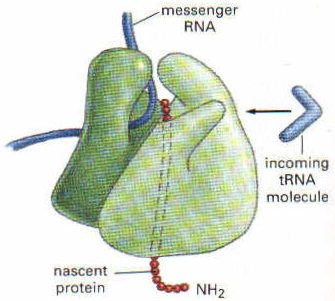
UAG, UGA ve UAA üç tane sonlanma kodonudur. Translasyon esnasında, ribozomun A bölgesi mRNA’daki bu sonlanma kodonlarından biri üzerine gelince peptid zincirinin sentezi durur. Bazı sonlanma faktörleri bu bölgeleri tanır ve buraya gelince hem mRNA’yı hem de proteini ribozomdan ayırır. Ortama serbestleşen polipeptid zinciri daha sonra bazı modifikasyonlara uğrayabilir.
Translasyon sonrası polipeptid zincirlerinde meydana gelen modifikasyonlar
Polipeptid zinciri sentezlenirken ya daha ribozomdan ayrılmadan önce ya da ayrıldıktan sonra, bazı amino asitler ayrılabilir (kısalma) veya bazı kimyasal gruplar polipeptid zincirine bağlanırlar (fosforilizasyon, glikozilasyon, hidroksilasyon, diğer kovalan bağlanmalar).
Bazı proteinler ilk sentezlendikleri zaman büyük, inaktif moleküller halindedirler. Endoproteaz enzimleri ile bazı kısımları kesilerek, daha kısa ama aktif hale geçerler. Bu kesilme işlemi endoplazmik retikulum, Golgi cismi veya sekretuvar veziküller içerisinde gerçekleşebilir. Proteinler ya hücre içindeyken kısalmaya uğrarlar ya da daha inaktif haldeyken hücreden çıkarlar ve etki gösterecekleri dokuya gittikten sonra burada kısalma işlemine maruz kalırlar. İnaktif halde salınan bu protein prekürsörlerine zimogen adı verilir.
Serin, treonin ve tirozin fosforilizasyona uğrayan amino asitlerdir. Fosforilizasyon olayı protein kinaz’lar tarafından katalizlenir. Fosfatazlar ise fosforilizasyonu bozarlar. Fosforilizasyon sonucu proteinin fonksiyonel aktivitesi artabilir veya azalır.
Hücre zarının yapısında bulunan veya dışına salgılanan proteinlerin çoğuna karbonhidratlar bağlanır. Karbonhidratlar serin, treonin ve asparajin amino asitlerine bağlanırlar.
Kollagenin yapısında bulunan prolin ve lizin amino asitleri hidroksillenirler.
DNA yapısında bozuklukların meydana gelmesi, bunların sonuçları ve teşhisi
RNA zinciri üzerinde bir nukleotid bazı değişirse buna nokta mutasyonu denilir. Bu durumda 3 sonuç gelişebilir. 1) Kodonun yapısı değişir, ama yeni oluşan kodon da aynı aminoasiti sentezler, neticede bir değişiklik olmaz (sessiz mutasyon); 2) Kodonun yapısı değişince sentezlediği aminoasit de değişir, neticede oluşan proteinin yapısı değişmiş olur, yani patolojik protein ortaya çıkar (yanlış mutasyon, missense mutasyon); 3) Baz değişikliği sonucu oluşan kodon, sonlanma kodonu haline gelirse, translasyon işlemi bu noktada kalır ve protein sentezi eksik olmuş olur (saçma mutasyon, nonsense mutasyon).
Mutasyon eğer önemli ise organizma ölebilir ya da hastalık durumları ortaya çıkar.
Genetik kod belirli bir başlangıç noktasından, nukleotid’den, itibaren okunmaya başlar ve kesintisiz olarak devam eder. Yani okunurken olduğu gibi üçlü gruplar halinde birbirlerinden ayrılmamışlardır. Aradan bir baz çıkar (delesyon) ya da fazladan bir baz eklenirse, bu noktadan sonraki kodonların içeriği de değişmiş olur ve sonuçta mutasyona uğrayan noktadan itibaren yanlış aminoasitler bir araya gelir, proteinin yapısı bozulur (çerçeve kayması mutasyon). Ama, 3 baz birden değişmiş yada eklenmişse bir çerçeve kayması olmaz, peptidden, bu üçlü nukleotid dizesinin kodladığı, sadece bir aminoasit eksilmiş veya eklenmiş olur.
Polimeraz Zincir Reaksiyonu (PCR= Polymerase chain reaction)
DNA zinciri üzerinde meydana gelen, nukleotidlere ait değişikliklerin saptanması için kullanılan bir laboratuvar yöntemidir.
Belli bir DNA dizesinin laboratuvarda çoğaltılması esasına dayanır. Böylece bir nukleotid dizesinden milyonlarcasını birkaç saat içerisinde çoğaltmak mümkün olmaktadır. Burada amaç, o DNA zinciri içerisinde aranılan bazların, ya da nukleotidlerin, eksilmiş olup olmadığını bulmaktır. Eğer tek bir DNA zinciri üzerinde bunu araştırmak istesek, materyal azlığından dolayı mümkün olmamaktadır. Veya klonlama ile vektörler içerisinde bu DNA kısımları çoğaltılmaya çalışılmakta olup, buda fazla zaman ve emek gerektirmektedir. PCR bu bakımdan son derece faydalı, pratik ve kolay bir yöntem olarak geniş çapta kabul görmüştür.
PCR tekniğinde, içinde eksik olup olmadığı (veya mutasyona uğrayıp uğramadığı ya da var olup olmadığı) araştırılacak DNA dizesi önce ısıtılarak denatüre edilir ve iki sarmal birbirinden ayrılır. Sonra ortama aranan nukleotid dizesini belirleyen primerler eklenir. Bu primerler her bir DNA zincirine tutunarak, onu kopyalarlar ve 2 zincirli yeni bir DNA sarmalı meydana getirirler. Arkasından bu DNA sarmalı da ısıtılarak iki zinciri birbirinden ayrılır, her zincire tekrar primer yapışır, tekrar DNA sarmalı sentezlenir, ve bu şekilde işlem tekrarlayarak, kısa süre içerisinde istenen nukleotid dizesinden bol miktarda elde edilir. Sonra bu nukleotid dizeleri jel elektroforez yöntemi ile ayrıştırılır. Eksik olan ya da aranan nukleotid dizesi burada kendini gösterir. Sonuçta aranan nukleotid dizesinin yokluğundan yani delesyonundan bahsedilebilir, veya varlığı gösterilmiş olunur.
DNA üzerinde araştırılan nukleotid dizeleri, ortama konulan sentetik primerler ile belirlenir. Bunun için aranan nukleotide ait her iki ucunda bulunan “kanat dizeleri”nin bilinmesi gerekir. Bunlar önceden bellidir. Yaklaşık 20-35 nukleotid uzunluğunda dizelerdir. Sentetik primerler bu dizelere komplementerdir. DNA sarmalı ısıtılarak birbirinden ayrıldıktan sonra, soğutulur. Bu sırada ortama konulan primerler, kendilerine ait kanat dizelerine gidip tutunurlar. O zaman ortama DNA polimeraz konularak, bu primerlerin yeni DNA zinciri sentezlemeleri başlatılır. Sentez için gerekli olan 4 çeşit deoksiribonukleotid trifosfatlar da ortama eklenilir. DNA polimeraz, primerin 3′ ucuna ortamdan aldığı nukleotidleri ekler. Ekleme işlemi hedef DNA boyunca devam eder.
Yeni oluşan her DNA çift sarmalı tekrar ısıtılarak denatüre edilir ve ayrılır, böylece oluşan yeni zincirlere tekrar primer bağlanır, yeniden DNA sentezlenir, bu işlem sürekli tekrarlanır. Birkaç saat içerisinde 20-30 kez bu döngü tekrarlanır. Sonuçta milyonlarca kopye elde edilmiş olunur.
PCR tekniğinin bir çok alanda yeri vardır. Örneğin mutasyonların veya delesyonların (Y kromozom delesyonunda olduğu gibi) gösterilmesi gibi. Adli tıpta da önemli faydaları bulunur. Erkek infertilitesinin tetkikinde PCR tekniği yaygın olarak kullnılmaktadır. Olguların %30’una yakınında Y kromozomu üzerinde spermatogenezden sorumlu bazı nukleotidlerin eksik olduğu bilinmektedir. İşte bu nukleotidlerin eksikliğinin (delesyonunun) ortaya konulması PCR ile yapılmaktadır. Ama çok sayıda sorumlu gen bulunması nedeniyle, her bir gen için ayrı bir primerin kullanılması gerekmektedir. Hangi genlerin araştırılması gerektiği konusunda henüz bir fikir birliği oluşmamıştır. Bu konuda çalışmalar halen devam etmektedir. Bazı genlerin delesyonunda testislerde hiç germ hücresi bulunmayacağı ya da belli bir evrede spermatogenezde duraklama olacağı ortaya konmuş olduğu için, bu olgularda TESE yapıp yapmamaya karar vermede, PCR ile o genlerin varlığının araştırılması çok önemlidir. Ayrıca, bu delesyona uğramış genleri içeren DNA’ların, doğacak çocuğa geçmesi durumunda aynı şekilde infertilitenin de doğacak çocukta görülme olasılığı bulunduğundan, üremeye yardımcı teknikler uygulanacak ailelerin bu konuda bilgilendirilmesi bakımından da, Y kromozom delesyonlarının tetkiki ayrıca önemlidir.
Faydalanılan Kaynaklar:
Alberts B: The Cell, Garland Pub, New York, 1994
Lewin B: Genes, Oxford University Press, Oxford, 1997
Champe PC, Harvey RA: Biochemistry, Lippincott, (çev. Tokullugil A ve ark, ed, Nobel, 1997) 1994
Passarge E: Color Atlas of Genetics, Thieme, New York, 1995