Prof.Dr. Kaan AYDOS
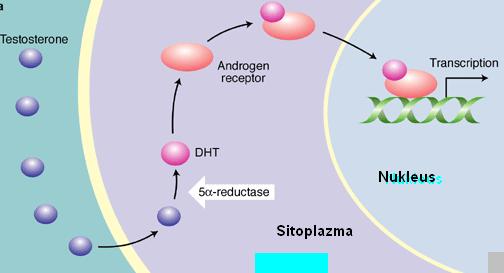
Dolaşımdaki esas androjen testosterondur. Testosteron kanda sadece %2 oranında serbest konumda bulunur. %60’ı steroid-bağlayıcı globulin (SHBG; veya testosteron-östradiol-bağlayıcı globulin, TeBG)’e, %40’ı ise serum albuminine, kortikosteroid-bağlayıcı globuline (transkortin), progesteron-bağlayıcı globuline ve alfa-asit glikoproteine bağlı durumda bulunur. Son 3 bağlayıcı proteine olan bağlanma çok düşük orandadır, esasen %60 SHBG ve %40 serum albumini olarak kabul edilebilir. Hedef organa ulaştığında testosteron SHBG’den ayrılır ve hücreye diffüzyonla girer. Organın özelliğine bağlı olarak testosteron hücre içerisinde DHT (dihidrotestosteron)’a dönüşür. Örneğin prostatta DHT aktif androjendir.
www-ermm.cbcu.cam.ac.uk/ 02005148h.htm
Androjen hücre içine girince önce androjen reseptörüne (AR) bağlanır. AR’leri glükokortikoiler, mineralokortikoidler, tiroid hormonu,retinol, östrojen ve progesteron gibi steroid hormon reseptörleri ailesine aittir. Mekanizmaları tipiktir: DNA’daki spesifik bölgelere bağlanırlar ve mRNA sentezini başlatırlar. Bu reseptörlerde N-terminal kısmı, DNA’ya bağlanan kısım ve hormona bağlanan kısım olmak üzere 3 bölüm vardır.
Diğer bölümleri fazla farklılık göstermemekle birlikte, N-terminal kısımları steroid reseptörleri arasında daha fazla farklılık gösterir. Bu ucun görevi tam anlaşılamamıştır. Reseptörün DNA’ya bağlanmasında veya transaktivasyonunda rol oynuyor olabilir. Ama içerdiği CAG dizeleri (glutamin aminoasidini kodlar) en önemli özelliğidir.
Androjen reseptörü bir yandan kendine spesifik geni aktive ederken, diğer yandan AR yapan geni de aktive eder (otoaktivasyon). Dolayısıyla, dokuda androjen seviyesi arttıkça AR seviyasi azalmaz, sabit kalır. İlginç olarak, bu sırada AR’ünü kodlayan mRNA seviyesi azalmaktadır. Bu tezat üzerinde araştırmalar devam etmektedir.
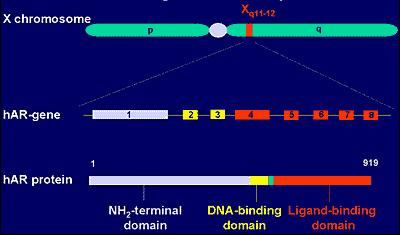
AR’nü kodlayan genler X kromozomu üzerinde lokalizedir. Parasentromerik bölgede uzun kol üzerinde q11-q13 aralığında bulunur. Erkekte sadece 1 tane X kromozomu bulunduğu için bu genden bir allel üzerinde lokalize sadece bir tek kopya mevcuttur.
www.endotext.org/male/ male3/male3.htm
AR’ünü kodlayan gen üzerinde 8 adet ekson vardır. Eksonlar genin AR proteinlerini sentez eden DNA kısımlarıdır. Bunlardan mRNA transkripsiyonu yapılır, mRNA’lar sayesinde de aminoasitler birleştirilerek protein sentezi için translasyon gerçekleşir.
AR‘ünün molekül ağırlığı 110 000 baz (yaklaşık 90 kb), içerdiği aminoasit sayısı ise 917’dir. Yukarıda tanımlandığı gibi 3 kısımdan oluşur: 1) Transkripsiyonu başlatan kısım (N-ucu): gendeki 1. ekson tarafından kodlanır, nukleustaki proteinlere ve gendeki regülatör ünitlere bağlanır; 2) DNA üzerinde hedef genlere bağlanan kısım (DNA’ya bağlanan kısım): ekson 2 ve 3 tarafından kodlanır; ve 3) C-ucundaki androjene bağlanan kısım: 4-8 eksonlarınca kodlanır.

Androjen reseptörü 3 bölümden oluşur: Transkripsiyonu başlatan kısım, DNA bağlanan kısım (DBD) ve androjen bağlayan kısım (LBD). Transkripsiyonu aktive eden kısım AR genindeki 1 numaralı exon tarafından, DBD 2 ve 3 numaralı, LBD ise 4-8 numaralı exonlar tarafından eksprese edilirler (herkules.oulu.fi/isbn9514267621/ html/x516.html)
AR geninin santral bölgesi yakınında bulunan genler, diğer bölgelerindeki genler ile etkileşime giren aminoasitlerin sentezini uyarır. Böylece AR’ünün AR geni üzerindeki diğer genlerin aktivasyonu da sağlanmış olunur.
AR’ünün DNA’ya bağlanan kısmı 72 aa’den meydana gelmiştir. Tüm reseptörün %10’unu oluşturur. N-ucu ile androjen bağlayan kısım arasında lokalizedir. Sisteinden zengindir. 12-13 aa’den oluşan iki adet protein lup oluşturur. Bu luplar bazalde çinko moleküllerine tutunurlar. Her bir çinko molekülüne 4 sistein (sülfür molekülleri ile) tutunduğu için toplam 8 sistein bu nedenle lup yapacak şekilde üstüne kıvrılır. Parmak şeklindeki bu luplara “zinc fingers” adı verilir. İşte bu luplar AR geni üzerindeki spesifik genlere bağlanırlar.
www.andrologysociety.com
İlk lup ekson 2 tarafından kodlanır ve esas DNA’ya bağlanan bölümü oluşturur. İkinci lup ise ekson 3 tarafından kodlanır ve DNA-reseptör etkileşimini stabilize etme görevi vardır. Çünkü ikinci lup bazik özellikte olup DNA’nın negatif yüklü fosfat gruplarıyla etkileşime girer. AR’ünün bağlandığı DNA kısmı androjenlere özel protein sentezlettirdikleri için ayrıca androjen responsive elements (ARE) adı da alırlar. Bu elementler genin uyarılması için emir verirler. Eğer ARE’e bağlanan aminoasit diziliminde bir mutasyon olmuşsa, reseptör androjene duyarlı genleri aktive edemez ve neticede androjene duyarsız kalıtsal sendromların ortaya çıkmasına neden olurlar.
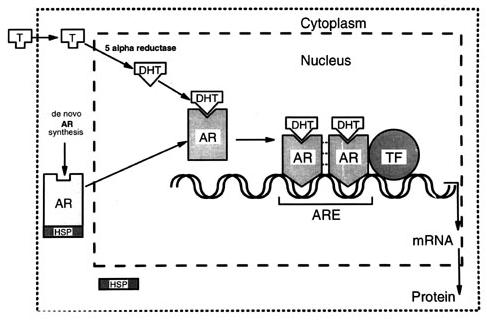
Androjen reseptörü DNA’ya birbirinin aynısı 2 hormon-reseptör kompleksinden oluşan bir homodimerik form formunda bağlanır. Dimerizasyon; ya androjenin reseptöre bağlandığı sırada ya da DNA’ya bağlanma sırasında gelişir. Sitozolden nukleusa geçen reseptör-hormon kompleksi, DNA üzerindeki kendine ait spesifik bölgeyi tanıyarak, oraya bağlanır. Daha önce belirtildiği gibi bu bağlanma reseptördeki 2 çinko parmaksı çıkıntıları ile DNA arasında gerçekleşir. Reseptörün dimerleri DNA üzerindeki ARE‘lere bağlanırlar.
www.endotext.org/male/ male3/male3.htm
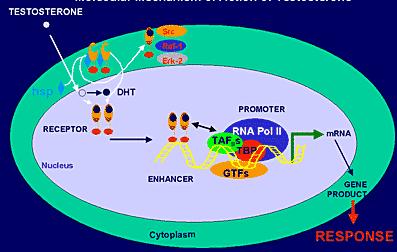
AR geninin DNA sekansları tekrarlayan sekanslar içeren (palindromik) tipik androjen-bağımlı gen özelliği taşır ve reseptörün tutunması için “bağlanma bölgesi” oluştururlar. Dimerik formdaki androjen reseptörünün DNA’ya bağlanabilmesi işte bu palindromik yapı sayesinde gerçekleşir. Ancak, bu palindromik yapının yanı sıra, palindromik yapıya benzemeyen sekansların bulunduğu DNA kısımları da mevcuttur. Androjen reseptörünün bu bölgelere bağlanması androjen+androjen reseptörü, RNA polimeraz, transkripsiyon faktörleri ve DNA’dan oluşan bir transkripsiyon kompleksinin oluşması ile sonuçlanır. Bu transkripsiyon kompleksi de androjen-bağımlı genlerden mRNA sentezlenmesini başlatır. mRNA’nın translayonunu takiben de androjen-bağımlı proteinler yapılır.
Transkripsiyonal kompleksin çalışması ko-aktivatörler ve ko-represörler tarafından kontrol edilir. Ko-aktivatörler reseptörün transkripsiyon fonksiyonunu artırırken, ko-represörler azaltırlar. Bunlardan bazıları : CPB/3000, ARA 54 ve testis spesifik ARIP-3’dür.
Reseptörün androjen bağlayan kısmı C-karboksil ucudur. Tüm reseptörün yaklaşık %30’unu oluşturur. Androjenlerin bağlanabileceği 24 aminoasitlik hidrofobik bir cep taşır. Testosteron ya da DHT’un bu bölgeye bağlanması ile reseptörün yapısı da değişir ve transkripsiyon yapıcı bir özellik kazanmasını sağlar, yani adeta bir transkripsiyon faktörü rolü üstlenmiş olur. Buna transaktivasyon denilir. Bu bölge, gen transkripsiyonunun kontrollü uyarılmasından sorumludur. Eksperimental olarak bu bölgenin çıkarılması durumunda, gen transkripsiyonu da artar. Zaten, “heatshock” proteinleri (örneğin HSP 90; protein 90) normalde bu bölge üzerindeki özel kısımlara bağlanarak reseptörün inaktif formda kalmasını sağlarlar. Normal şartlarda sitozolde reseptör serbest konumda dolaşır. Testosteron reseptöre bağlanınca, reseptörde bazı yapısal değişimler ortaya çıkar. Bunun sonucunda HSP reseptörden ayrılarak fonksiyonel domainlerin serbestleşmesi sağlanmış olur. İşte bundan sonra hormon-reseptör kompleksi nukleusa taşınabilir, dimerize olur ve DNA’ya bağlanabilir.
Reseptörün bir başka özelliği ise, sentezlendikten sonra fosforilizasyona uğramasıdır (post-translasyon yapılanma süreci). Fosforilizasyon reseptöre dayanıklılık sağlar.
DHT testosterona göre daha fazla afinite ile bağlanır. Çünkü testosteron reseptörden daha çabuk ayrılmaktadır. Androstenadion, östradiol ve progesteron gibi diğer androjenler ise reseptöre testosterona göre daha düşük afiniteye sahiptirler.
Bazen AR’ündeki C-karboksil ucuna ait mutasyonlar reseptörün fonksiyonunu artırır, yani daha aktif olur veya uygun olmayan steroidlerce de aktif hale gelir bir özellik kazanır ve DNA’ya bağlanarak hedef genini uyarır, buradanda yeni AR’ü sentezlenir. Sonuçta reseptörün bu bölgesindeki aminoasitler çıkarılsa da, uyardığı DNA tarafından androjen reseptör proteini sentezlenebilir ama böyle reseptör proteinleri artık androjen bağlayamazlar. Androjen bağlayamazlar ama başka steroidlerce uyarıldıklarında, androjenler ortamda olmasa bile, transaktivasyon yapabilirler. Bunun önemi fonksiyonel testlerde transkripsiyonun olduğu izlenimi vermeleri ama sentezlenen AR proteininin artık androjen bağlayamamasıdır. Prostat kanserinde AR mutasyonu gelişerek östrojen hatta antiandrojenler gibi ilgisiz steroidler böyle reseptörleri aktifleştirebilirler. İşte prostat kanserinin androjen duyarsız hale gelmesinin ya da androjenler olmasa bile, ürettikleri yeni AR’leri ile (androjenler olmaksızın diğer steroidlerce uyarılabilen) prostatın büyümesinin nasıl devam ettiğinin yanıtı budur.
AR’ünün nukleusa taşınabilmesi için androjen bağlanması gerekir. Ama nukleusa girdikten sonra reseptörün DNA’ya bağlanması için artık androjen gerekmez.
AR’ünün transkripsiyondan sorumlu kısmı (N-domain) nukleustaki bağlanmada önemlidir. Burada aminoasitlerin belli bir sırada dizilmiş olmaları gerekir: 20 glutamin-8 prolin-23 glisin ünitinden oluşur. Glutamin dizeleri gendeki CAG dizeleri tarafından kodlanarak sentezlenir. Gen üzerinde normalde 11-31 (bazılarına göre 17-29) adet CAG dizesi bulunur. Sayıdaki bu değişkenliğe polimorfizim denilir. Dolayısıyla, erkekler arasında AR’ünde bulunan poliglutamin dizelerindeki sayı da farklılık gösterir. Farklı etnik gruplar arasındaki prostat kanseri sıklığındaki değişkenlik buna bağlıdır.
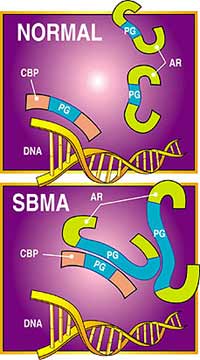
Androjen reseptörü (AR) üzerindeki glutamin dizelerindeki artış (AR genindeki CAG dizeleri tarafından kodlanır) sonucunda gen regüle edici protein CBP’yi DNA’dan ayırarak kendine bağlanmasına yol açar. Neticede, CBP’nin kontrol ettiği DNA kısmı normal fonksiyonunu göremez ve spinal-bulbar muscular atrophy (SBMA) ya da Kennedy hastalığını ortaya çıkarır.
CAG tekrarlarının sayısı azaldıkça, prostat kanserinin metastaz sıklığı da artar. Kennedy hastalığında ise (X-bağımlı spinal ve bulbar muskuler atrofi ile karakterize olup motor nöronları etkiler), glutamin tekrar sayısı 40-60’a (bazılarına göre 72’ye) kadar artmıştır. Dize sayısı arttıkça koaktivatörlerin etkisi ve neticede transkripsiyon da azalır. İşte bu nedenle Kennedy hastalığında androjen duyarsızlığı da görülmektedir. Glutamin dizeleri arttıkça azoo- ya da oligozoospermi ortaya çıkar. Bu nedenle androjen insensitivitesi veya aşırı virilizasyon durumları gibi kalıtsal özellik taşıyan hastalıklarda AR yapısının araştırılması önem kazanmaktadır.
Embriyonda AR’nün gelişmesi önce ventral prostat ve seminal veziküllerin mezenşiminde görülür. Daha sonra epitel hücrelerinde de gelişmeye başlar. Seminal veziküllerin ve Wolff kanalının glandüler gelişiminde primer androjen testosterondur. Ürogenital sinüsten prostat gelişiminde ise DHT etkendir. Testosteron ve DHT’un her ikisi de AR’üne bağlanabilirler ama DHT 3-10 kat daha potenttir.
Çok sayıda modülatör proteinler AR’üne bağlanarak doku spesifisitelerini, DNA’ya bağlanabilme potansiyelini ve nukleusa taşınabilme özelliklerini etkileyebilirler. Androjen reseptörleri birbirleriyle birleşerek homodimerleri, ya da bu regülatör proteinlerle birleşerek heterodimerleri oluşturur.
Androjen reseptörleri vücutta hep aynı değişmez yapıda bulunduklarına göre, reseptörün dokuya göe davranışının ne olacağını belirleyen faktörler nelerdir? Aynı hayvanın farklı dokularında bulunan iki hücre tipi nukleuslarında aynı AR’lerini içeriyor olabilir. Ama bir dokuda bu hücreler AR’üne farklı yanıt vererek farklı protein sentezlerlerken, diğer dokuda aynı tip hücre AR’ünün uyarmasına farklı bir protein yaparak yanıt verebilir. Burada dokuya özgü ko-aktivatörler ve ko-represörler sorumludurlar. Bunlara selektif androjen reseptör modülatörleri (SARMs) denilir. Aynı selektif östrojen reseptör modülatörlerinde olduğu gibi, SARMs de örneğin sadece kas gelişmesini etkileyip kardiyovasküler risk faktörlerini etkilememeleri gibi selektif bir etki göstermektedirler. Örneğin hem prostatta hem de seminal veziküllerde AR bulunur ve ortamda DHT varsa DHT her iki dokuda da aynı genoma bağlanır ama prostatın androjen kontrolündeki gen aktivasyonu neticesi seminal veziküllerinkinden farklı bir gen ekspresyonu oluşarak değişik bir protein sentezlenmektedir. Aynı DNA içeren iki hücrenin aynı reseptöre neden farklı yanıtlar verdiği henüz izah edilmiş değildir. Burada reseptörler ultrastrüktürel yapıda farklılık gösteriyor olabilirler ya da bazı DNA dizeleri metilasyona uğrayarak veya kromatin yapısı bakımından modifiye olmuş olabilirler. Bu anlamda androjen reseptörleri sitozol içerisindeyken bir sitozolik aktivatör gibi davranmış olur. Örneğin DHT’un mitojen-aktive edici protein kinazları uyararak hücre çoğalmasını kontrol etmesi gibi.
Androjen reseptörü tarafından aktive olan genlerin yaptıkları ürünlerin neler olduğu tam izah edilmiş değildir. Bunlardan sentezlenen proteinler aynı zamanda diğer moleküler olayları da başlatmaktadırlar. Primer ürünlerin uyarması ile sentezlenmiş sekonder proteinler arasında probasin, faktör IX ve PSA sayılabilir. PSA’yı sentezleyen gen aktivasyonu androjen etkisi altındadır. Çünkü PSA geni bir “androjen bağlayıcı” eleman içerir. Bu eleman da androjen reseptörü ile bağlanmaktadır. Androjenlerin aynı zamanda PSA üzerinde posttranskripsiyonel etkileri de bulunur. Androjenler bir yandan gen transkripsiyonunu başlatırlarken, diğer yandan gen sentezini inhibe de edebilmektedirler.
Delesyon ve inaktive edici mutasyonlar gibi androjen reseptör bozuklukları reseptör fonksiyonunu etkiler. Sonuçta fenotipte virilizasyon azalmasından testiküler feminizasyona kadar değişen farklılıklar ortaya çıkar.
Androjen reseptör geninin tamamen ortadan kalkması, androjen aktivitesindeki komple eksiklik nedeniyle kadın fenotipinin ortaya çıkmasına neden olur. DNA-bağlayıcı ya da androjen-bağlayıcı kısımların mutasyonları gibi androjen reseptör fonksiyonlarını ciddi şekilde bozan durumlarda da benzer klinik tablo görülür. Oysa N-bağlayıcı kısımdaki mutasyonlarda androjen reseptör fonksiyonları tamamen ortadan kalkmamakta sadece azalmakta olduğu için, idiyopatik erkek infertilitesinde rol oynar. CAG tripletlerinin sayısı reseptörün transkripsiyon aktivitesini değiştirir ama spermatogenezi ne derecede etkilediği tam izah edilememiştir. Bazı oligozoospermik erkeklerde hormon bağlayan kısımda mutasyonlar saptanmıştır. Dimerizasyon bozuklukları transaktivasyonu bozmakta, buda ko-aktivatörlerin bağlanmasını ve sağlıklı transkripsiyonel komplekslerin oluşmasını engellemektedir. Prostat kanserli hastalarda da androjen reseptör mutasyonları gösterilmiştir. Bu mutasyonların tümör gelişimine katkıları tartışmalıdır.
Androjen reseptör mutasyonları ile fenotip arasında doğrudan bir ilişkinin gösterilememiş olması, androjen fonksiyonlarında reseptörün yanı sıra başka faktörlerin önemine de işaret etmektedir.
KAYNAKLAR
Campbell’s Urology, 7th ed, Walsh, Retik, Vaughan, Wein (eds), WB Saunders Co, Philadelphia, 1998
Andrology: Male Reproductive Halth and Dysfunction, 2nd ed, Nieschlag E, Behre HM (eds), Springer, New York, 2000